O colesterol é derivado da Acetil CoA Citossólica
A maioria das células animais são capazes de sintetizar colesterol,
a formação de colesterol só e substancialmente ativa nas células
hepáticas. A função das lipoproteínas em parte esta associada a
veiculação do colesterol à dieta e derivado do fígado para o restante
das células do corpo.
A descoberta de que todos carbonos provêm do Acetil CoA usando
marcadores radioativos, foi o principio da descoberta da síntese de
colesterol. Outro fato foi a descoberta que o intermediário na
biossíntese do Colesterol é o esqualeno e que é formado a partir do
Isopreno.
Acetato (C2) ---> Isoprenóide (C5) ---> Esqualeno(C30) ---> Colesterol (C27)
Estrutura do isopreno
O colesterol é sintetizado a partir da acetil CoA citossólica, que é
transportada da mitocôndria pelo sistema de transporte do citrato,
sendo também usada na síntese de ácidos graxos é metabólito importante na
ramificação da via da biossíntese de lipídios.
A condensação sequencial de 3 moléculas de Acetil CoA ,com formação do
intermediário HMG CoA (Hidroximetilglutaril CoA) levando a formação do
Mevalonato, é o primeiro estágio na formação do colesterol e as etapas
de condensação são catalisadas por isoenzimas citossólicas de enzimas
mitocondriais envolvidas na formação de corpos cetônicos.
Reações iniciais da biossíntese de colesterol levando à formação de
m evalonato
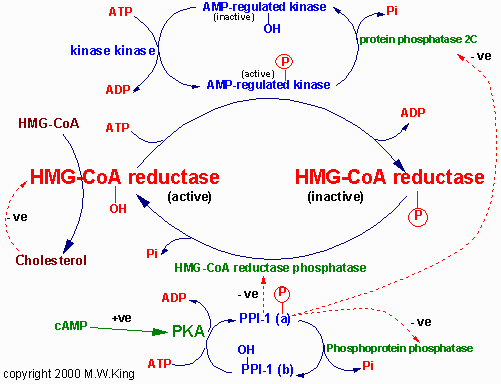
A HMG CoA redutase, interconversível e inativada por fosforilação,
catalisa a redução do HMG CoA a Mevalonato limita a velocidade da via
global da síntese de Colesterol. A Insulina aumenta a atividade da
redutase e o Glucagon tem efeito inverso, embora as etapas entre
recepção hormonal e o estado de fosforilação permanecem
desconhecidos.
Moléculas de Acil CoA de cadeia longa causam inibição da enzima
podendo
levar a efeito alostérico direto na enzima ou de efeito de cinase que
catalisa a fosforilação e a inibição resultante da HMG CoA redutase. A
atividade de HMG CoA redutase também é controlada pela concentração de
colesterol, maiores concentrações levam a formação de derivados do
colesterol que inibem a enzima alostericamente. Os altos níveis de
derivados levam a um aumento de degradação e diminuição da síntese da
enzima.
Veja como a HMG-COA redutase é regulada
O Mevalonato é convertido a Isopentenil pirofosfato em várias etapas
catalisadas por enzimas resultando na sua fosforilação, uma segunda
fosforilação e descarboxilação.
Formação de isoprenil pirofosfato
Uma isomerase catalisa a conversão do isopentenil pirofosfato a seu
isômero, diametilalil pirofosfato, que se condensa, cabeça - cauda,
como isopentenil pirofosfato formando o geranil pirofosfato. A
condensação de 2 moléculas em C5 para formação de um ionte carbônio, intermediário
da dimetilalil pirofosfato, que é atacado pelo isopentenil
pirofosfato.
Esta reação é repetida: C10 , geranil pirofosfato condensa cabeça com
cauda com o o iospentenil pirofosfato formando farnesil pirofosfato,
C15. duas moléculas de farnesil condensam-se do mesmo modo formando o
esqualeno, C30.
Um intermediário, lanosterol, acumula-se em grandes quantidades em
células ativando a síntese de Colesterol. As etapas entre esqualeno e
lanosterol envolve adição de oxigênio seguida de ciclização da cadeia
formando um núcleo esteróide com quatro anéis. A conversão do
lanosterol ao colesterol ocorre por via de múltiplas etapas envolvendo
deslocamento de metilas, oxidações e descarboxilações.
Veja um esquema da sintese do colesterol
Diversidade de produtos relacionados ao colesterol e a via metabólica
de síntese de colesterol
O metabolismo do colesterol é a fonte de um grande número de outros
constituintes celulares. O isopentenil pirofosfato é precursor de um
grande número de produtos, como vitaminas lipossolúveis A, E e K e a
ubiquinona em células animais, e terpenos, plastoquinona e fitol,
cadeia lateral da clorofila, em vegetais.
O colesterol é precursor de sais biliares que facilitam a digestão de
gorduras; vitamina D que estimula a captação de Ca 2+ pelos
intestinos;hormônios esteróides tais como testorona, estradiol e esteróides
que controlam o equilíbrio salino. O principal produto da síntese de
esteróis é o próprio Colesterol, que modula a fluidez da membrana e é
um componente essencial da membrana citoplasmática das células animais.
Transporte do colesterol.
O colesterol e outros lipídios orgânicos são transportados no sangue,
para locais específicos, por lipoproteínas. O triacilgliceróis
exportados pelo intestino são carreados por quilomícrons e
hidrolizados por lipases das paredes dos capilares dos tecidos alvo. O
Triacilgliceróis são exportados do fígado por VLDL (lipoproteína de
muito baixa densidade), que depois de liberar o seu conteúdo é
convertida em IDL (Lipoproteína de densidade intermediária), e
depois em LDL (Lipoproteína de baixa densidade ). A IDL e LDL carreiam
esteres de colesterol (principalmente linoleato de colesterila), a LDL e
captada por células Hepáticas e de tecidos periféricos por endocitose
mediada por receptor de LDL por receptor. O receptor de LDL, atravessa
a membrana plasmática, liga a LDL e participa da sua entrada. A ausência
do receptor na forma homozigotica de hipercolesterolemia familiar leva
a um aumento de LDL- colesterol, deposição de colesterol na paredes de
vasos sangüíneos e ataques cardíacos em crianças.
Veja um esquema de uma lipoproteína do plasma
Veja uma animação super interessante de como que as lipoproteínas de transporte viajam pelo sangue clicando aqui.
Pode demorar um pouco para carregar mas vale a pena!!!
{kind=link}
{kind=link}