RESUMO: Os autômatos são entidades que reproduzem determinados comportamentos dos seres vivos. Na Idade Média eram comuns os autômatos mecânicos e atualmente eles se tornaram tambem sistemas lógicos inteiramente abstratos ou de uma grande complexidade eletrônica, como os computadores. Inspirados nesses autômatos muitas tentativas de explicação de funcionamento dos seres vivos foram feitas. Por outro lado, admite-se hoje que todos os mecanismos biológicos devam basear-se em princípios físico-químicos, que vão aos poucos sendo demonstrados. Esses achados vão se integrando ao enorme edifício do nosso conhecimento e fazendo ampliar as fronteiras de nosso questionamento. Assim, o modelo é uma representação do real que descrevemos com certas margens de erro ou de que simulamos deliberadamente só algumas de suas propriedades. A primeira etapa do modelo biológico é apenas um esquema com alguma semelhança ao objeto. A seguir ele pode adequar-se a mesmo identificar à certa dinâmica do objeto, tornando-se uma lei de seu funcionamento. Há um maior amadurecimento do modelo ao obter uma equivalência entre os símbolos do modelo e as partes do objeto. Finalmente o modelo é submetido a um teste de múltiplas variáveis para ver sua maior capacidade de ajuste aos resultados experimentais. Nessa progressão há uma maior generalização, através de um trabalho de constante retorno às origens, reiniciando com um constructo mais complexo, o que lhe confere maiores graus de liberdade e maior capacidade de adaptação, quando técnicas adequadas são empregadas, tais como as cibernéticas, as probabilisticas, as matemáticas e as de simulação.
HISTÓRICO
Durante a Idade Média, com o desenvolvimento das corporações artesanais, era usual a associação de maravilhosos relógios, mecânicos ou hidráulicos, com certos movimentos de animais e bonecos. Essa tradição chegou até nós através dos relógios-cuco e dos famosos relógios públicos de cidades europeias como Estrasburgo e Munique. É desse tempo também as figuras de macaco dos relógios islamitas. Elas eram famosas por apresentar movimentos tão semelhantes ao humano. Assim, é compreensível a atitude de Santo Tomás de Aquino em afirmar, na Summa Theológica, "que os animais exibem comportamento regular e ordenado, cabendo, pois, encará-/os em termos de máquinas, distinguindo-se eles do homem por haver sido dotado de alma racional, atuando, conseqüentemente, por força da razão" (27). Sob forte influência espiritualista, Descartes conseguiu um compromisso entre a explicação do corpo humano pelos fenômenos fisicos e a alma. Mas, carecendo de base experimental e baseado em analogias, ele explicava o movimento dos olhos pela pressão hidráulica do músculo do lado oposto. Hoje sabemos que os olhos são puxados pelos músculos e não empurrados. Essa concepção foi importante para colocar também o corpo humano sujeito a explicação de mecanismos que a experimentação buscaria comprovar.
No século XVII já foi possível construir as primeiras máquinas de efetuar cálculos matemáticos e notáveis autômatos musicais, o que constituía uma ousada incursão na simulação do próprio comportamento racional do homem. Não tardou haver uma série de reações para delimitar o terreno das simulações biológicas (4, 23). No século XIX, já sob influência do racionalismo crescente, apareceram outros ramos da ciência que explicavam a evolução das espécies e lançavam as bases de um materialismo total. No início do século seguinte floresciam grupos que explicavam o comportamento como um condicionamento e, em contracorrente ao racionalismo, o mesmo comportamento era atribuido a instintos básicos. Essa última interpretação coincidiu também com um amplo movimento em defesa dos princípios básicos do indeterminismo na física (10, 45). A perplexidade das considerações diante do fenômeno vital levaram o positivismo original e suas explicações a naufragarem.
Nos meados do século XX houve um grande avanço nos mecanismos automáticos de controle de trabalhos, nas técnicas de transmissão, codificação e decodificação de mensagens, nos processos lógicos de um raciocínio e na sintaxe da transcrição de um pensamento. Assim, ressurgiu um novo racionalismo, fortalecido pelo grande avanço da matemática, da lógica, da estatística, da teoria das probabilidades, da eletrônica e da tecnologia em geral. Novos campos da ciência são lançados cada vez mais na interface das ciências da vida; a cibernética, em 1948, a teoria da comunicação, em 1949, a biônica e uma nova matemática mais apropriada a certas características biológicas está sendo desenvolvida nessa última década (teoria da catástrofe). Esse neopositivismo vai permitir grandes avanços nas ciências da vida, como atestam os numerosos modelos de funcionamento de partes do sistema nervoso (16, 24, 25, 26, 32, 33, 34, 38), apoiados geralmente por uma aperfeiçoada infra-estrutura de coleta e medida de dados. Inúmeras outras proposições são feitas do lado das ciências exatas (2, 20, 30, 32, 33) e do lado das ciências biológicas (8, 32, 39, 43). Em geral o sucesso foi encontrado nos modelos de partes periféricas do sistema nervoso e células isoladas. Também houve preferência para se trabalhar em animais simples como polvo, aplisia, caranguejo, camarão e peixe. Os progressos na neurofisiologia deram também grandes detalhes aos funcionamentos das funções sensoriais e das atividades dos membros e dos músculos, relacionados com funções centrais. Houve grande redução da barreira que separava o cérebro e seus mecanismos físico-químicos- Por outro lado, com a construção de fantásticos computadores, substituindo mesmo o raciocínio, como nos casos de ajuda aos diagnósticos médicos (36) os homens mergulharam numa manipulação criativa, profunda, lógica e controlada de simbolos. Todas essas abstrações eram propriedades dos cérebros, daí o nome cérebros eletrônicos (14). Vários autores passaram a recomendar em certos casos o abandono da causalidade restrita para uma generalização moderna (28, 42).
Nessa onda de otimismo, pretendeu-se construir um modelo de todo o cérebro com elementos lógicos de computação. Mas, já para representar um simples neurônio, grandes concessões foram feitas para evitar excessivas complexidades. O projeto ficou reduzido a simulação de algumas propriedades de um circuito nervoso, que significa extrema simplificação do objetivo inicial (35). Muitos textos de divulgação apareceram, mas carecem de uma metodologia mais confiável, fazendo generalizações indevidas, quando não apresentavam meros esquemas, cultuando o aparecimento de uma nova panacéia: a técnica e um novo racionalismo. Diante desses e outros fatos podemos compreender as ponderações de Covian (6): "O homem de ciência está se tornando cauteloso, prudente e não tão satisfeito dentro do limitado conhecimento da realidade. O biólogo moderno recusa as explicações universais realizadas por uma generalização ingênua a partir do setor parcial que intelectualmente domina''. Há assim uma atitude de prudência comum entre os neurofisiologistas, mesmo entre os mais otimistas (7, 21).
OS MODELOS BIOLÓGICOS NA ERA E DA CIBERNÉTICA E O APARECIMENTO NA TEORIA DOS AUTÔMATOS
Richalet et al. (33) classificam assim as etapas da elaboração de um modelo em geral:
1 . Caracterização
2. Adequação
3. Identificação
4. Reificação
5. Verificação
Os fisiologistas, tradicionalmente, quando bem sucedidos, dão por encerrados seus trabalhos na etapa 3, depois de traçarem curvas, detetarem tendências, acharem correlações, calcularem coeficientes e expoentes, o que não difere muito de outros ramos da ciência (42). Eles seguem ainda hoje uma abordagem geralmente analítica, muitas vezes beneficiados por uma técnica moderna e mais refinada. O experimentador espera coletar bastantes dados para deduzir de sua amostragem o comportamento de todo o sistema. A análise unitária, entretanto, além das dificuldades, sobretudo em se tratando do sistema nervoso central, pode ser influenciada por sistemas de retroalimentação ou de ativação celular, que dependem muito mais de propriedades de numerosos circuitos nervosos e de complexas reações bioquímicas. As abordagens modernas com enfoques sistêmicos são capazes de solucionar essas dificuldades. É verdade também que a análise de sistemas depende de um conhecimento preciso do funcionamento dos componentes isoladamente (38). Por outro lado, a simples inclusão de noções de teoria de circuitos para encontrar analogia entre os sistemas levaram muitos autores a atingir apenas a etapa 1, porque existem muitos tipos diferentes de mecanismos com a mesma relação entre entrada e saida, sem apresentarem, entretanto, correspondência entre as partes (29).
É evidente que, diante da complexidade do ser biológico e cedendo a tantas exigências do modelo, visualizado de uma maneira intransigente, isto é, não se admitindo uma margem de erro, só podemos chegar a um impasse, a saber, ou o modelo do objeto é o próprio ser natural, ou temos um caso inteiramente abstrato, sem ligação com os seres naturais, como são os isomorfismos da matemática moderna (3, 15). Assim o modelo factivel na ciência aplicada é a representação aproximada da estrutura e das propriedades de um sistema material, trazendo consigo a noção do erro conhecido, proveniente da comparação e das medidas de cada variável. Ai reside sua grande vantagem, pois, uma vez conhecidos também os limites, o modelo poderá ser o veículo que expresse bem o objeto de análise atraves da nossa linguagem. Entretanto, os trabalhos, apresentados sem a devida prudência, não atendendo a uma completa satisfação das etapas de construção de um modelo sem margem de erro conhecida ou satisfazendo somente certos aspectos do desempenho biológico, provocaram com justa razão uma série de críticas dos biologistas e modelistas. Em conseqüência, criou-se um espírito de incompreensão, que foi importante para Shannon e McCarthy (37) resolverem dar nome a um novo campo da ciência, a teoria dos autômatos, para trabalharem com mais liberdade teórica e menos restrições da comparação direta com o sistema biológico. Essas entidades, que satisfaziam sobretudo aos itens caracterização e adequação, eram livres das contingências de reificação e de verificação, especialmente sob a ótica de múltiplos estímulos. Esse campo expandiu-se rapidamente com apoio de muitas teorias matemáticas modernas e obteve um desenvolvimento espetacular nas ciências da computação (1, 12, 40, 41). O interessante é notar que várias propriedades dos seres vivos foram retidas tais como: movimento próprio, reprodução de outro sistema idêntico e a si mesmo, aprendizagem, adaptação, sensibilidade, capacidade de auto-reparação e de perseguir um objetivo, memória, processamento de informação e resposta conseqüente, desenvolvendo com novas abordagens uma outra espécie de biologia puramente teórica (16, 18, 27, 31, 37).
ALGUNS PASSOS NA DIREÇÃO DOS MODELOS BIOLÓGICOS
As abordagens aos modelos biológicos podem ser resumidas nas quatro grandes classes seguintes: pelos circuitos eletrônicos, pelas formulações matemáticas, pelas simulações de computadores e pelas bases tônicas (15). Vamos examinar o exemplo de um caso simples em que o tratamento foi uma formulação matemática mesclada com noções de teoria de circuitos usando elegante artifício. Durante muitos anos foi aceita a lei de Fechner, que descreve a relação entre a magnitude do efeito da estimulação subjetiva (E) e da intensidade dos estímulos (I) colocada na seguinte fórmula:
|
K = constante |
Entretanto trabalhos mais recentes de Stevens, Mountcastle e seus colaboradores demonstraram que a lei realmente seria da forma:
|
(1) |
|
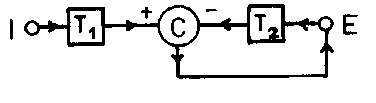
MacKay (22) esquematizou tal função num circuito com um transformador logarítmico (T1), dando entrada num comparador (C), onde simultaneamente se associava outro transformador logarítmico (T2), vindo da saida do comparador em forma de uma retroalimentação negativa (Fig.1). Ele conseguiu ter uma resposta exponencial na saida do comparador. Matematicamente isso seria o correspondente ao seguinte:
T1 log I - T2 log E = 0 , ou
E = IT1/T2 (2)
Fig. 1. Esquema de um circuito com retroalimetação negativa que adapta a lei de Stevens às variáveis: intensidade dos estímulos (I) na entrada e a magnitude do efeito da estimulação subjetiva (E) na saida. T1 e T2 são respectivamente transformadores da forma K1.log e K2.log das funções I e E, e C é um comparador (modificado de MacKay (22) ).
Assim, nesse tratamento simples, podem ser observadas as duas primeiras etapas de Richalet (33) numa aproximação a um modelo de percepção. Entretanto, a terceira etapa, a identificação dos parâmetros está comprometida. A constante "a" da fórmula (1) só pode ser igual a um. A adequação do expoente pode levar ou 1o ) ao seu cálculo numérico, adaptado a certo teste em particular ou 2o.) a demonstrar a necessidade de se procurar outro modelo para os outros casos. De toda maneira a etapa de reificação estaria muito comprometida, uma vez que ninguém ainda imaginou um sistema físico ou químico capaz de tal desempenho (29). Para uma melhor evolução posterior desses modelos é necessário partir de um sistema com um grande desenvolvimento experimental que elucide profundamente as propriedades da atividade receptora em cada modalidade de sensação em particular, como estão encaminhados por exemplo os trabalhos do grupo de Mountcastle (44).
Por outro lado, partindo de uma função de transferência com parâmetros a determinar, haveria mais liberdade de se ajustar o modelo com as propriedades do ser natural. O resultado dessa abordagem pela teoria dos circuitos é visto na Fig. 2, onde as respostas de um fuso muscular ao estiramento súbito, medidas pelo inverso das freqüências de potenciais de ação, é comparado com as respostas simuladas no modelo (34). É tão grande a semelhança das respostas do modelo e do objeto que mesmo um experimentador especializado não distingue um do outro. Depois, no próximo passo podemos identificar as variaveis do fuso muscular com os parâmetros da função de transferência, decorrente do modelo:
|
H (t) = ----------------------------- (3) |
|
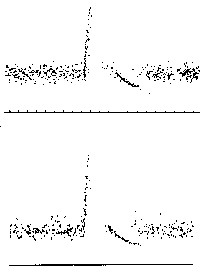
Fig. 2. Comparação das respostas de um fuso muscular ao estiramento súbito (acima) com a simulação de um modelo (abaixo). Nas ordenadas o inverso da freqüência instantânea e nas abscissas o tempo (Modificado de Richalet et al. (34)).
Os parâmetros dessa outra equação 3 são suscetíveis de se adequarem a outros dados experimentais, tais como as constantes de tempo e duração de contração, tornando-se, portanto, explícitas. Mas a seguir, várias perguntas ainda ficaram sem respostas como a) qual a resposta de contração a uma estimulação em forma de degrau e b) a contração é local ou propagada. Novas reconsiderações obrigam a abandonar a hipótese tomada inicialmente de um sistema com constantes localizadas, se quisermos também satisfazer a um teste mais complexo de verificação. Isso vai favorecer também encontrar um significado biológico mais amplo com maior número de parâmetros utilizados. No entanto, serão necessários novos dados experimentais que dependem praticamente do progresso das pesquisas do grupo de Laporte, para comprovação desses outros desempenhos (34). Assim, para elaboração de um modelo mais avançado devemos retornar ao inicio com outra função de transferência com mais variaveis para ter maiores possibilidades de obter melhor reificação e mais ampla verificação do modelo. Essa nova função não contradiria a primeira, mas a deveria engloba-la como um caso particular no momento de fazermos uma redução de variaveis que nos leve de volta as variaveis anteriores.
Em suma, nesses últimos decênios houve uma grande proliferação e progressão qualitativa dos modelos biológicos. Uns mais especulativos, partindo de propósito de grandes simplificações e dando ênfase a certos fatos bem definidos como os mais relevantes. À medida do necessário se introduziriam também outros parâmetros importantes. A maior critica a esse método seria o maior afastamento da realidade, tornando a identificação muitas vezes impossível de se atingir. O outro método reproduz com grande fidelidade as propriedades neurais, apresentando relativa identidade com o objeto, guardando uma margem de erro bem estabelecida, mas arcando com mais complexidade para evoluir. Uma das maiores contribuições dos modelos são as indagações novas que eles suscitam, permitindo novas montagens experimentais para a elucidação de muitas facetas desconhecidas do desempenho biológico e em alguns casos permitindo mesmo a previsão de algumas respostas.
REFERÊNCIAS
![]()