| <<voltar |
PESQUISA > Peixe elétrico, caramujo e mamíferos > Link 11 > |
Departamento de Ictiologia , INPA, MANAUS, Universidades da California em San Diego (a), EUA, Federal de Minas Gerais (b), Brasil, e Gottingen (c), RFA, e Techniches Hoschule, Darmstadt (d), RFA.
RESUMO: Após fazer-se um estudo sobre aspectos gerais de famílias, gêneros e algumas espécies de Gymnotoidei e outros peixes elétricos amazônicos, bem como sobre seus habitats e hábitos de vida, analisaram-se os órgãos elétricos e respectivas descargas em várias espécies. Procuram estabelecer-se as funções da descarga do órgão elétrico (DOE) na vida do animal, correlacionando-se a mesma com o comportamento e outros eventos. Em seguida eestuda-se a eletrorrecepção, seus órgãos dos sentidos especiais e processamento cerebral, inclusive o uso de eletrorrepção por peixes não elétricos. Conclçui-se o trabalho, interpretando-se o possível significado ecológico e prático dos fatos observados.
SUMMARY: Aspects of the use of electric organ discharge and electroreception in amazonian gymnotoidei and other fishes. An overview of the families, genera and some species of the Gymnotoidei is presented with a review of their behaviour and natural history, Characteristics of the electric organs and electric organ discharge (EOD) for various species are presented. The role of EOD on the animal's life is discussed, especially as it relates to behaviour and other events. Electroreception, its special sense organs and brain-processing, induding use by non-electric fish is discussed. The results of these studies are interpreted in terms of their pratical and ecological significance in the life of this group of fish.
I) Introdução ao estudo de peixes
elétricos
A bacia amazônica é o ambiente natural
de um grande e notável grupo de espécies de peixes, os Gymnotoidei,
os quais possuem, todos, órgãos elétrícos.
Além da conhecida enguia elétríca, muitas outras espécies
semelhantes dentre incontáveis milhões de peixes, estão
descarregando fracos pulsos elétricos na água, noite e dia,
durante toda a vida, numa.frequência de poucos até mais de
mil pulsos por segundo.
O Electrophorus electricus, a enguia legendária,
desperta um especial interesse. Como os outros peixes fortemente elétricos,
o "catfish" do Nilo, Malapterurus e a raia elétrico marinha,
Torpedo - os quais pertencem a dois grupos não relacionados
- sua estranha capacidade de excitar tanto a sensação quanto
a contração foi percebida, senão compreendida, muito
antes de Benjamim Franklin ter empinado seu papagaio durante uma tempestade.
O poder da enguia elétrica tem despertado tanto respeito que, entre
os caboclos que vivem ao longo dos rios do Amazonas, ainda existe a crença
de que sua descarga ao redor da base de uma palmeira pode fazer com que
as frutas caiam ().
lgualmente notáveis são as outras
espécies de gimnotóides com nomes comuns, como sarapó,
ituí, tovira, peixe espada e fantasma negro. A descarga de seus
órgãos elétricos (daqui para frente apenas DOE) é
demasiado fraca para ser sentida, muito menos para poder ser ofensiva ou
defensiva, e é usada para dois outros propósitos - para a
detecção de objetos- alimento, plantas, obstáculos,
cavidades ou de outra espécie, Estas funções requerem
um sistema nervoso altamente desenvolvido para a percepção
de mudanças muito pequenas no fraco campo elétrico da DOE
e também para o controle da DOE, pulso por pulso.
A capacidade para detectar campos elétricos
extremamente fracos, na água, não está confinada nas
poucas famílias que possuem órgãos elétricos
e geram DOEs. Vários grandes grupos de peixes - embora seja minoria
entre todos os peixes - são receptores mas não emissores,
pelo menos de pulsos não especializados.Todos os peixes, como de
fato todos os seres vivos, dissipam alguma corrente elétrica, ao
menos como uma espécie de escapamento dos processos internos, e
este pequeniníssimo sinal é usado pelos peixes eletrorreceptivos
para encontrar a presa. No Amazonas, nove famílias de Siluroidei
(em inglês "catfish", no Brasil surubim, cuiú-cuiú,
caparari, bagre, piraíba, etc.) mais os Elasmobranchia residentes
e visitantes (tubarões e raias) possuem delicados eletrossensores
na pele.
Este trabalho tem por objetivo introduzir um aspecto
raro mas significativo de comportamento e ecologia. É uma atraente
pesquisa em prática na Alemanha, na Holanda, na Inglaterra, na França,
nos Estados Unidos, na Rússia e em outros locais, inclusive no Brasil.
Cientistas que vivem perto dos habitats naturais do peixe elétrico
têm oportunidades únicas para aplicar estas especializações
a problemas gerais da biologia de peixes e .águas naturais.
II) ASPECTOS GERAIS DE FAMÍLIAS, GÊNEROS
E ALGUMAS ESPÉCIES DE PEIXES ELÉTRICOS
A. GYMNOTODEI
Todos os peixes elétricos de água
doce do Novo Mundo pertencem a esta subordem da ordem Cypriniformes e aparentemente
todos os membros da subordem são elétricos. Para o propósito
de nosso trabalho, é importante dizer que aparentemente todas estas
espécies também possuem eletrorreceptores - a julgar pela
amostra das espécies, que têm sido estudadas e que são
algumas em cada uma das famílias conhecidas. Todos os gimnotóides
são delgados e alongados, sem nadadeira dorsal mas com uma nadadeira
anal extraordinariamente longa, lembrando a lâmina de uma faca pontuda
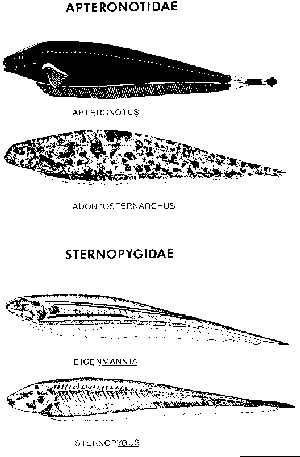
daí o nome de "peixe-espada" (Fig. 1). Uma família tem a
forma de uma enguia.
Seis famílias são reconhecidas pelas
mais recentes autoridades (Mago-Lecia, 1976), conforme relação
abaixo, com alguns de seus gêneros representativos:
1. Electrophoridae. A enguia-elétrica, compreendendo o gênero Electrophorus (poraquê), forma uma família própria. É o maior gimnotóide, o único com alta voltagem, capaz portanto de usar a descarga para ataque ofensivo ou defensivo. Possui também órgãos elétricos separados de baixa voltagem, que emitem pulsos numa baixa frequência, irregularmente, para eletrolocalização e eletrocomunicação.
2. Gymnotoidae. O gênero comum Gymnotus (sarapó ou carapó) é um grupo euritópico, largamente distribuído, de espécies muito semelhantes. G. carapo tem sido muito estudado. É um peixe de pulsos (definido mais abaixo) de frequência média (cerca de 50 por segundo) e pode atingir mais de 45 cm, alimenta-se não apenas de invertebrados mas também de peixes de vários centímetros de comprimento.
3. Rhamphichthydae. Este grupo de peixes de freqüência média inclui o gênero Rhamphichthys, que atinge mais de um metro de comprimento, e Gymnorhamphichthys, o mais nítidamente circadiano de um grupo geralmente noturno. Ambos os gêneros normalmente ficam imóveis sobre o fundo, durante o período de repouso, sem procurar manter uma postura com o dorso para cima.
4. Hypopomidae. Este é um grupo mais diversificado de peixes de freqüência de pulso; baixa (<30 por segundo), média e alta (>l00 por segundo), incluindo muitas espécies de Hypopomus, Hypopygus, Parupygus e Steatogenys. Existe uma acentuada diferença entre as espécies na taxa de repetição e uma tendência para correlacionar inversamente este fato com a duração do pulso; permitindo presumivelmente que, espécies de pulsos simpátricos, reconheçam os sinais específicos de sua própria espécie.
5. Sternopygidae. Esta é a primeira família de espécies produtoras de ondas (em oposição a pulsos, no capítulo IV). Sternopygus é um gênero de baixa frequência (70-140 por segundo), portanto com o pulso mais longo da subordem (acima de 7 ms). Eigenmannia de várias espécies têm uma descarga de menos de 200 a mais de 500 por segundo, e são bons exemplos de espécies que ficam suspensos na água, com contínuas ondulações da nadadeira anal. Uma onda caudal e outra rostral encontrando-se e cancelando-se na parte mediana do peixe. Este gênero também é um bom exemplo de grupo gregário, já que a maioria dos outros gimnotóides é territorial ou mantém espaçamentos entre si.
6. Apteronotidae. Estes são os peixes de alta freqüência, produtores de ondas (cerca de 700-2100 por segundo), notáveis também por possuirem os ritmos mais regulares, em repouso, conhecidos no mundo vivo. As pequenas modulações de freqüência, sob controle cerebral, possuem significado comunicativo. Gêneros comuns na Amazônia incluem Apteronotus, (antigamente Sternarchus), Sternarchella, Sternarchorhynchus, Sternarchorhamphus, Sternarchogton e Porotergus.
B. OUTROS PEIXES ELETRORRECEPTIVOS
Órgãos dos sentidos especializados
na detecção de fracos campos elétricos na água
têm sido "inventados" pela evolução muitas vezes -
e não apenas nos vários grupos não relacionados de
peixes que desenvolveram órgãos elétricos (Rajidae,
Torpedinae dos Elasmobranchia; a família
Fig. 1 - Peixes gimnotóides. São mostrados exemplos de 4 gêneros em 2 famílias comuns no sistema do Amazonas
Astroscopidae de teleósteos marinhos; os teleósteos
africanos de água doce dos Mormyrldae, Gymnarchidae e Malapteruridae,
os teleósteos de água doce da América tropical da
superfamília Gymnotoidei), mas também em outros. Estes incluem
todos os outros Elasmobranchia (tubarões e raias), principalmente
marinhos, mas também as raias de água doce, Potamotrygon
e Paratrygon da América do Sul. Entre os peixes ósseos,
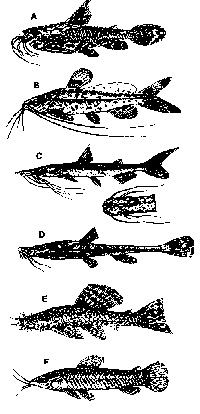
o maior grupo eletrorreceptivo é o dos Siluroidei (Fig. 2), uma
superfamília que abrange mais de uma dúzia de famílias,
incluindo nove na América do Sul o milhares de espécies.
De acordo com Bates (1964), a fauna extraordinariamente rica de peixes
da América do Sul (i.e. a lista das espécies) é 40%
de siluróides. Alguns dos mals comuns ou melhor conhecidos são
os Pimelodidae, incluindo Sorubim, Pseudoplatystoma (capararí)
e Brachiplatystoma (piraíba), os Doradidae, incluindo Doras
e Oxydoras (cuiú-cuiú), os Callichthyidae, incluindo
Corydoras, os Loricariidae (cascudos), incluindo Ancistrus
(barbadinhos) e Plecostomus, os Trichomycteridae (Pygidiidae), incluindo
Vandellia (candiru).
Entre outros peixes, todos da América do
Sul e outras partes, existem evidências, sugestivas ou conclusivas,
de que a eletrorrecepção também ocorre nos Chondrostei,
incluindo Scaphirhunchus (esturjão), Polyodon ("spoonbill"),
Dipnoi, incluindo Lepidosiren (pirambóia, peixe pulmonado e possivelmente
mesmo em alguns anfíbios da ordem Gymnophiona (=Apoda, Caeciliidae,
cobra de duas cabeças).
A eletrorrecepção não é
conhecida em invertebrados ou em outros vertebrados aquáticos. Em
vertebrados terrestres, incluindo a espécie humana, há alguma
razão para crer que, mesmo se nenhum orgão de sentido está
presente para detectar campos elétricos aplicados externamente,
pode ser normal algumas células do sistema nervoso serem influenciadas
por fracas correntes no tecido. Portanto, os mecanismos que estudamos nos
estranhos peixes podem ter um mais amplo interesse neurobiológico.
Fig. 2 - Peixes siluróides. São mostrados exemplos de várias famílias de "catfishes" comuns no sistema do Amazonas. A. DORADIDAE, Doras (Acanthodoras) spinosissimus. B. PIMELODIDAE, Rhamdia sebae e C. Sorubim lima (com vista dorsal da cabeça). D. BUNOCEPHALlDAE, Bunocephalus bicolor. E. LORICARIIDAE, Ancistrus cirrhosus. F. CALLICHTHYIDAE, Callichthys callichthys (de Sterba, 1963).
A maioria das espécies está sujeita
a grandes mudanças sazonais, com a subida e a descida nos níveis
da água, a inundação de enormes áreas de florestas
e mudanças drásticas em tipos e disponibilidade de alimento.
É provável que muitas espécies
sejam especialistas alimentares, por exemplo buscando alimento ao redor
de folhas de plantas aquáticas ou raízes de plantas flutuantes,
ou entre o folhedo do fundo ou no lodo. A julgar pela experiência
com os grupos mais bem conhecidos, pode esperar-se que especialistas extremos
em alimentos e em associados e inimigos não sejam raros entre os
gimnotóides. Todos os gimnotóides parecem ser carnívoros
e a maioria alimenta-se de insetos e crustáceos, embora as espécies
maiores predem peixes - talvez incluindo outros gimnotóides. Electrophorus
e Gymnotus apanham e consomem peixes elétricos pequenos em
tanques, mas isto não nos auxilia muito na estimativa do grau de
uma tal predação na natureza.
Sabemos menos ainda a respelto de outros inimigos
dos gimnotóides e presumívelmente eles devem precisar escapar
de um grande grupo de predadores, como os siluróides. Em várias
localidades, métodos de coleta que apanham todos os peixes em vários
metros cúbicos de água rasa [cerca de 1 m de profundidade),
trazem um grande número de indivíduos e a maioria deles pode
ser gimnotóides. Isto poderia dizer que o grupo possui significado
econômico como uma parte substancial da biomassa disponível
para os membros mais altos da cadeia alimentar.
Fig. 3 - Órgãos elétricos. São mostrados esquematicamente exemplos das células geradoras de corrente e sua inervação em 2 tipos muito diferentes de órgãos elétricos, ambos dos gimnotóides. A. A enguia eÍétrica com seus órgãos de alta e baixa voltagem; o esquema em maior aumento mostra os compartimentos do tecido conectivo organizado serialmente, cada um contendo uma camada de eletrócitos com uma especialização de suas superfícies rostral e caudal, de tal modo que apenas uma é despolarizada pela chegada do impulso nervoso. Cada uma destas membranas pode contribuir com mais de 0,1 V (de Altamirano et al., 19530. B. O ituí-cavalo, uma espécie onduladora de alta freqüência e baixa voltagem; seu órgão consiste de terminações nervosas especializadas dos axônios efetuadores. Os nervos repetidos a cada segmento vertebral, orientados na mesma direção adicionam a corrente de seus grandes nódulos de Ranvier (de Bennett, 1971).
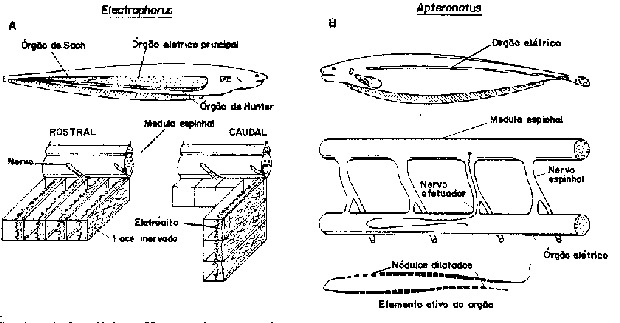
Órgãos miogênicos são
capazes de produzir descargas mais poderosas de que os órgãos
neurogênicos. Na enguia-elétrica (Eletrophorus), o
órgão miogênico pode descarregar rapidamente (1 ms)
uma corrente de 1 ampère numa voltagem de 500 volts (Fig. 3A).
A vantagem dos órgãos neurogênícos
é que eles podem descarregar mais freqüentemente do que os
órgãos miogênicos. Isto decorre especialmente devido
à capacidade dos neurônios de descarregar em taxas mais altas
do que as células musculares. Certas espécies dos apteronotídeos
descarregam o orgão elétrico a mais de 1800 vezes por segundo.
Fazem isto, dia e noite, durante toda a vida.
Os eletrócitos miogênicos estão,
geralmente, entre as maiores células do animal e podem ser em forma
de fita, disco ou fuso. Freqüentemente estão dispostos em pilhas,
com todas as células orientadas no mesmo sentido. Esta disposição
é comum na maioria dos órgãos miogênicos e parece
ser um resultado da evolução convergente para permitir uma
produção máxima de voltagem pelo órgão.
A enguia-elétrica possui três órgãos
elétricos: o de Sach, de Hunter e o órgão principal.
O órgão de Sach e a porção posterior do órgão
de Hunter podem ser descarregados sem uma descarga acompanhante do orgão
principal. Esta descarga é de baixa amplitude (10 V) e provavelmente
é usada mais para a detecção da presa do que para
a sua captura. A notória descarga de 500 volts é produzida
quando todos os órgãos descarregam simultaneamente; a maior
contribuição é do orgão principal. A maior
parte da corrente produzida pela descarga de alta voltagem é canalizada
diretamenie para o ambiente, reduzindo assim o efeito sobre os tecidos
do próprio animal, mas não será surpreendente descobrir
que o sistema nervoso central do animal possui adaptações
especiais que o tornam tolerante às correntes residuais que devam
fluir através dele .
Os eletrócitos ("eletroplacas") do órgão
principal na enguia-elétrica são em forma de fita, comprimidas
rostro-caudalmente e se estendem lateralmente desde a linha mediana. No
adulto existem cerca de 6.000 eletroplacas dispostas em série, em
uma coluna rostro-cau.dal. Existem cerca de 25 dessas colunas dispostas
em paralelo para formar o orgão principal. Como a maioria dos outros
eletrócitos miogênicos, estes são fisiologicamente
polarizados. As duas faces dos eletrócitos diferem, em termos de
sua excitabilidade elétrica. A face posterior dos eletrócitos
da enguia é inervada, eletricamente ativa e produz um pico de potencial
elétrico devido ao sódio em resposta ao sinal de comando
neural. A face anterior possui uma baixa resistência e é eletricamente
inexcitável. Combinada com tecidos acessórios isolantes,
esta polarização maximiza a produção da corrente
elétrica. Parcialmente, devido a esta propriedade de polarização
do eletrócito, o estudo dos vários tipos de órgãos
elétricos conduziu a importantes reve!ações a respeito
das propriedades das células excitáveis.
Em céluIas musculares normais, a contração
é iniciada por um rápido evento elétrico que se proçaga
através da célula. Numa célula única, a amplitude
de pico a pico deste evento elétrico é de 80 a 100 mV. Este
evento, o potencial de ação do músculo é gerado
como um resultado de uma estimulação sináptica proveniente
de um neurônio que traz informação do sistema nervoso
central. A maioria dos eletrôcitos, embora não todos, assim
como as células musculares das quais são derivados, produzem
um potencial de ação tudo ou nada quando excitados pela informação
carregada pelo neurônio inervador. Com os eletrócitos dispostos
em série, como pilhas em uma lanterna, e estimulados simultaneamente,
as voltagens das descargas dos eletrócitos individuais se somam.
Se não ocorre curto circuito das descargas, as 6.000 eletroplacas
dispostas serialmente na enguia-elétrica adulta produzirão
uma descarga elétrica somada de 480 a 600 Volts, o que de fato ocorre.
Fig. 4 - Tipos de descargas de órgãos elétricos. São mostrados vários exemplos de gimnotoides da América do Sul para ilustrar a variedade de taxas de repetição e formas de ondas. As primeiras 4 espécies pertencem ao grupo chamado de "espécies pulsadoras", porque o intervalo entre as descargas é relativamente longo, As últimas 3 espécies pertencem ao grupo chamado de "espécies onduladoras", pois o intervalo é aproximadamente igual à duração da descarga. As DOEs são mostradas simultaneamente em escalas de tempo mais lentas ou mais rápidas (de Hagiwara & Morita, 1963).
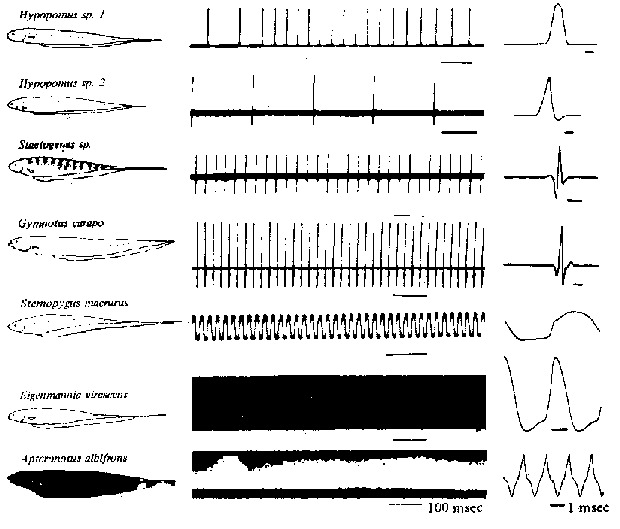
Com base na descarga de seus órgãos
elétricos, os peixes elétricos são classificados como
pulsadores ou onduladores (Fig. 4). Os pulsadores produzem uma descarga
curta semelhante a um pulso, a intervalos relativamente longos e irregulares.
Dependendo da espécie, um pulsador descarregará seu orgão
elétrico desde poucas vezes por minuto até mais de 80 por
segundo. Os onduladores descarregam numa freqüência constante
para produzir um campo elétrico de tipo semelhante à onda
senoidal. A regularidade dos intervalos de descarga é notável;
é suficientemente estável para ser o mais acurado dos relógios
biológicos. Dependendo da espécie, um ondulador descarregará
cerca de 100 vezes por segundo até mais de 1.800 por segundo.
Os pulsadores possuem uma considerável amplitude
para variar a taxa de descarga. Quando eles estão ativos, alimentando-se,
perturbados, ou de outro modo excitados, eles podem aumentar a taxa de
descarga em várias centenas por cento. Uma razão plausível
para este aumento dessa taxa associada ao estimulo, é que o peixe
interroga seu ambiente mais freqüentemente para detectar objetos e
dessa forma obter a informação necessária à
sobrevivência.
Os onduladores normalmente não alteram a
taxa de descarga em mais do que cerca de 10%, e apenas sob circunstâncias
particulares (ver RES em C do capitulo seguinte). O ondulador constantemente
obtém informações sobre o ambiente numa alta taxa,
.e as mudanças na freqüência das descargas estão
associadas mais estreitamente com comunicação social do que
com detecção de objetos.
O órgão elétrico descarrega
em resposta a um sinal de comando do sistema nervoso central. Este sinal
origina-se em um núcleo de células localizado na porção
do cérebro chamada bulbo e, com exceção do "catfish"
elétrico, é transmitido à medula espinhal por outros
neurônios. No "catfish" elétrico existem apenas duas células
neurais de comando na medula espinhal, e cada uma delas se ramifica profusamente
para inervar todos os eletrócitos de cada lado do corpo.
O orgão elétrico descarrega uma vez
para, cada sinal de comando. Na enguia-elétrica, há exceção,
pois uma série de sinais do comando de baixa freqüência
produz a descarga elétrica fraca. O desencadeamento de um comando
de alta freqüência causa potenciais de junção
sub-limiares nos eletrócitos do órgão principal e
na parte anterior do órgão de Hunter; estes potenciais imediatamente
somam-se para produzir uma descarga de alta voltagem, a qual então
segue os sinais de comando na proporção de 1 : 1.
Uma descarga máxima será produzida
quando todos os eletrócitos descarregarem simultaneamente. Isto
significa que o sinal de comando deve efetivamente chegar aos vários
eletrócitos simultaneamente. Existem dois modos principais pelos
quais a descarga sincrônica de eletrócitos distribuídos
espacialmente pode efetuar-se: a) pelo comprimento constante do caminho
neuronal dos núcleos de comando aos eletrócitos, com um caminho
direto aos eletrócitos distais a um caminho em circuito aos eletrócitos
proximais, e b) pe!a gradação nos tempos de condução,
com uma condução mais rápida do núcleo de comando
aos eletrócítos distais do que aos proximais.
Na enguia-elétrica, a sincronia é
efetuada pelos tempos de condução das fibras inervadoras.
As fibras que inervam os eletrócitos anteriores (fisicamente mais
próximos ao núcleo de comando) possuem menor diâmetro
e velocidade de condução mais baixa do que as fibras que
inervam os eletrócitos mais posteriores.
Fig. 5 - Eletrolocação. A. Um peixe elétrico, flutuando entre dois bastões de plexiglas suspensos verticalmente (secção transversal á esquerda), segue um movimento sinusoidal, S (t) e S (t) + D, por translação lateral do corpo F [(t). Esta resposta é prejudicada e ocorrem freqüentes colisões quanto menores forem os diâmetros F dos bastões escolhidos. Todos os bastões são escolhidos dentro do molde da agar para cancelar as diferenças nas informações mecânicas. Uma placa vertical de plexiglas em frente ao animal que flutua estabiliza sua posição entre os bastões oscilantes ([diagrama à esquerda). As análises de Fourier da resposta de seguimento do animal: F (t), revelam um forte componente na freqúência dos estímulos, S (t), que é de 0,2 Hz em cada registro. A amplitude deste componente, relativa à amplitude de S (t), é chamada "ganho", sua diferença da fase em relação à fase S (t) de "fase" nas últimas figuras (de Helligenberg, 1974). B. "Ganho" e "fase" da resposta de seguimento de Eigenmannia como uma função do diâmetro do bastão de plexiglas, numa frequência fixa de oscilação de 0,1 Hz e distâncla interna entre os bastões de 6 cm. O animal flutuou no centro do conjunto oscilante e portanto estava aproximmadamente a 3 cm de cada bastão. Símbolos diferentes representam valores médios para espécimes diferentes, as barras verticais representam desvios-padrões. Valores médios e desvios-padrões foram calculados a partir de distribuições bidimensionais de dados originais, representados no plano complexo, com ganho e fase como raio e ângulo, respectivamente (colocados mais acima, à direita). Desvio-padrão do ganho é definido como o desvio na direção radial, o desvio-padrão da fase é definido como perpendicular ao último (de Heiligenberg, 1973).
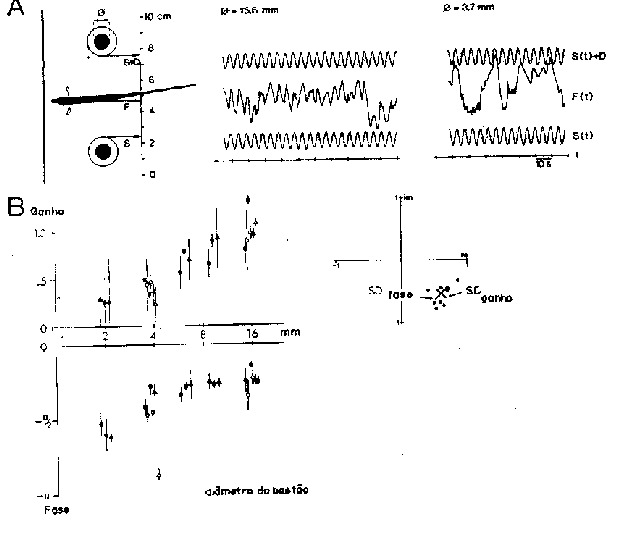
A capacidade ativa de eletrolocalizar objetos em
seu ambiente e passiva -de detectar sinais elétricos fracos, permite
ao peixe elétrico, como os gimnotóides, viver em águas
de pouca visibilidade, ser mais ativos durante a noite e permanecer escondidos
durante o dia. Desta forma, eles minimizam a captação por
predadores visuais, como ciclídeos e siluróides.
A eletrolocação ativa baseia-se no
seguinte princípio. A corrente da descarga do orgão elétrico
flui sucessivamente através da pele, da água que circunda
o animal e volta através da pele. Os.eletrorreceptores estão
localizados em poros na superfície do corpo e agem como monitores
das intensidades locais do fluxo da corrente transcutânea. Objetos
próximos da superfície do corpo do animal intensificarão
ou atenuarão o fluxo da corrente transcutânea local, conforme
sua resistividade seja mais baixa ou mais alta do que a da água
circundante. Estas mudanças locais nos padrões de fluxo representam
a "imagem elétrica" dos objetos. A intensidade e o contraste de
tais imagens são uma função da resistência relativa
da pele e geometria do corpo. Por exemplo, um longo filamento caudal, típico
para muitas espécies de peixe elétrico, amplia a faixa de
eletrolocação do animal e o contraste das imagens elétricas
é aumentado conforme o animal curve seu corpo e cauda ao redor dos
objetos de interesse.
As imagens elétricas são projeções
bidimensionais do ambiente sobre a superfície do corpo do animal,
o que pode ser então comparado com a retina no domínio da
visão. Os eletrorreceptores projetam-se somatotopicamente em camadas
nas unidades de ordem mais alta no lobo da linha lateral, torus semicirculares
e cerebelo (ver mais abaixo). lnterações mútuas entre
unidades adjacentes são acionadas para o processamento da imagem
elétrica e detecção da forma. Os eletrorreceptores
são assim organizados em padrões de campos receptivos, os
quais assemelham-se aos campos receptivos visuais em termos de centros
excitatórios, vizinhanças inibidoras, sensibilidade a movimentos
unidirecionais etc. Parece que a visão e a eletrolocação
desenvolveram princípios semelhantes no processamento de imagens
bidimensionais do ambiente.
A eletrolocação assemelha-se à
ecolocação no sentido em que o animal avalia seu ambiente
por uma retroalimentação continuamente avaliada por suas
próprias ações do que por uma exploração
passiva de fontes de energia estranhas, como a luz solar. Necessitamos
de novas pesquisas sobre os parâmetros usados na eletrolocação
em várias espécies, os limites de utilidade em relação,
por exemplo, com a forma, tamanho, posição, movimento, contraste
da água em condutância óhmica e a presença e
geometria da reatância capacitativa.
Fig. 6 - Eletrocomunicação. Estão ilustrados uns poucos exemplos de modulações de freqüência da DOE de Gymnotus carapo, associadas com o comportamento agonístico, A-C. Alguns padrões motores agonísticos. A. Disposição lateral antiparalela. B, serpenteamento mútuo (as setas indicam as curvas dos corpos dos peixes), C. Disposicão frontal mútua. D, E. Atividade elétrica concomitante. D. Freqüência instantânea das DOEs representada contra o tempo. Exibições indicadas para o peixe n0. 1 (D) e n0. 2 (E). (de Black-Cleworth, 1970).
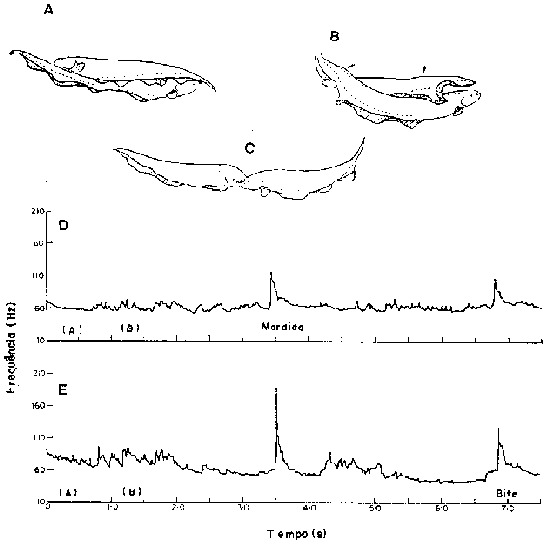
As diferenças entre espécies quanto
às características espectrais e temporais de DOE e a seletividade
associada dos eletrorreceptores às DOEs das espécies, certamente
aumentam o reconhecimento e a separação das espécies.
Espécies simpátricas estreitamente relacionadas podem geralmente
ser identificadas mais prontamente por suas DOEs do que pelas clássicas
características morfológicas. As colorações
do corpo servem primeiramente para a camuflagem do animal, não possuem
nenhuma função de aviso na comunicacão intra-específica
e são muito variáveis. As características de DOE,
provavelmente como conseqüência de seu papel na comunicação
social, apresentam considerável variabilidade entre espécies,
mas têm uma constância surpreendente dentro de cada espécie
(Fig. 4). Representam mais do que o canto do grilo como informações
seguras para a identificação das espécies.
Fig. 7 - A Resposta de Esquiva Saltando (RES) em Eigenmannia. A. Um peixe tentando escapar de estímulo que simula um vizinho, exceto que ele muda sinusoidalmente sua freqüência. Os deslocamentos de esquiva do peixe têm uma complexa seqüência no tempo dependendo da faixa de variação particular da frequência do estímulo (aqui cerca de 380 - 408 Hz), sua modulação de freqüência (aqui cerca de um ciclo em 20 segundos) e da amplitude e orientação do seu gradiente de voltagem. O estimulo efetivo é provavelmente a dlferença de frequência (D F=F estímulo - F peixe) e isto está continuamente mudando. B. Um método estudado da RES enquanto se mantêm o estímulo (D F constante). Um circuito que corrige automaticamente a diferença de frequência monitora as mudanças do peixe na freqüência e controla o estímulo para manter uma diferença constante de cerca de +4 Hz por 25 segundo, depois -4 Hz por 25 segundo e assim por diante. O peixe tenta aumentar a diferença mas não desloca mais do que uns poucos Hz antes de fixar-se num platô; isto depende da voltagem e orientação do estimulo. C. Os componentes principais deste comportamento (de Bullock et al., 1972 a, b).
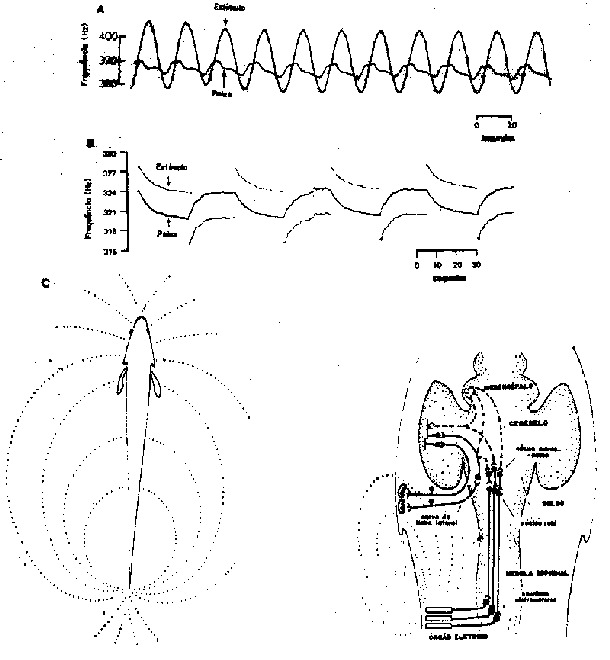
Espécies onduladoras enfrentam um problema
diferente, visto que toda outra descarga em forma de onda irá interagir
com a DOE do peixe para formar um batimento numa frequência diferente
(D F). Para manter a capacidade de eletrolocação, aparentemente
é suficiente manter o D F acima de cerca de 10 Hz. A RES nas espécies
onduladoras varia desde uma. tendência simétrica: para aumentar
a freqüência de DOE na presença de um baixo -D F e diminuir
a frequência de DOE na presença de um baixo + D F, até
uma ênfase assimétrica sobre uma dessas duas mudanças,
negligenciando a outra (Fig. 7). A maioria dos individuos de Eigenmannia
tendem a ser simétricos. Apteronotus responde tipicamente
apenas ao -D F, deslocando a freqüência para cima; depois retorna
em resposta ao + D F ou para a remoção do estímulo.
A RES é graduada com a intensidade do estímulo (normalmente
a proximidade do vizinho) e com o D F; é máxima em um D F
de cerca de 4 Hz, tanto em Eigenmannia quanto em Apteronotus.
Possui um teto e um mínimo, i.e., o peixe não se afasta muito
de sua frequência normal de DOE; geralmente a alteração
máxima é de 5 a 15 Hz. A constante de tempo é lenta;
após um rápido início de 0,15-0,2 segundos (o que
é tão rápido quanto o reflexo humano de afastamento
de um estímulo doloroso), a RES total é atingida apenas após
algumas dezenas de segundos, Daí em diante, ela pode ser mantida
tonicamente na presença do D F. O peixe não foge espacialmente,
i.e., move-se para longe da proximidade dos vizinhos em associação
com a RES. Algumas espécies são territoriais e espaçam-se
mutuamente mesmo que suas freqüências não sejam próximas.
Outras são gregárias e permanecem juntas mesmo que suas freqüências
sejam muito próximas. Desde que a variação natural
da taxa da DOE nas espécies onduladoras estende-se sobre várias
centenas de Hz, e apenas vizinhos em cerca de 10 Hz causem uma RES, a maioria
dos peixes não induz RES uns nos outros. Sternopygus, curiosamente,
parece não ter uma RES distinta.
Fig. 8 - A Resposta Ventral ao Substrato (RVS). Eigenmannia virescens mostra mudanças de posição em resposta a objetos no ambiente. Este comportamento garante uma orientação do corpo na qual o lado ventral do mesmo fica mais próximo ao substrato quando o peixe nada perto dele. A RVS é guiada pelo sentido elétrico e assim permanece após a cegueira. Os dados seguintes foram obtidos destes espécimes, A) Enquanto flutua acima de uma placa de plexiglas inclinado de 45°, um Engenmannia cego exibe uma inclinação média de 30.° (a ) em relação ao eixo de rotação (RVSr). B) Esta resposta de inclinação é fortemente reduzida se a placa de plexiglas é substituida por uma placa de agar mais ou menos transparente eletricamente. As RVSs em relação ao eixo de inclinação (C inclinação para cima; D inclinação para baixo) são mostradas na parte inferior da figura. As medidas foram tomadas para a posição da cabeça (b ) e da cauda (g ) tanto na situação de inclinação para cima (RVSc) quanto na de inclinação para baixo (RVSb). Os dados numéricos foram baseados em 8 espécimes (de Meyer, original).
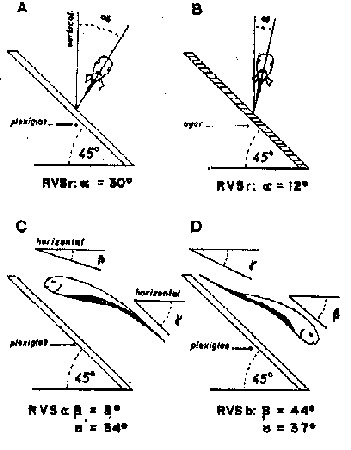
Além da RVS em relação ao eixo
de rotação, outras respostas também ocorrem em relação
ao eixo de inclinação (Fig. 8C e D). Se o peixe estiver orientado
com seu plano sagital na vertical e perpendicular à placa de plexiglas
de 45°, observa-se uma VSR no plano vertical. Se o peixe estiver flutuando
acima da placa, com a cabeça voltada para ela, isso resulta numa
resposta de inclinação para cima (RVSc). As medidas dos desvios
da cabeça e cauda da horizontal são muito diferentes (Fig.
8C). Quando orientado na direção contrária à
placa, o peixe exibe uma resposta de inclinação para baixo
(RVSb). Neste caso, os ajustes da cabeça e causa são semelhantes
(Fig. 8D) . Novamente os desvios da horizontal são muito menores
com uma placa de agar, mais ou menos transparente eletricamente.
Estas observações demonstraram que
a posição de flutuação de Eigenmannia
depende da orientação do fundo ou do objeto que está
próximo ao peixe. Contudo permanece obscuro o porquê destes
animais terem desenvolvido um tal mecanismo de controle de posição.
É provável que as observações no campo possam
esclarecer um pouco mais a respeito do significado deste padrão
de comportamento para Eigenmannia em seu habitat normal. Tem sido
sugerido que a RVS de certos peixes de recife, guiada visualmente, facilitaria
a camuflagem e a obtenção de alimento do substrato. Possivelmente
vantagens do mesmo tipo resultam deste comportamento em Eigenmannia.
TABELA 1 - Gimnotóides encontrados na região de Manaus, Amazonas, e sua taxa de repetição de descarga do orgão elétrico (DOE). As espécies são grupadas de acordo com a forma de onda da descarga como ou "espécies pulsadoras" ou "espécies onduladoras" (ver capítulo V para definições). Medidas feitas por W. Heiligenberg and J. Bastian em Novembro, 1976, senão estiver especificado. Identificações supostas.
| ESPÉCIES PULSADORAS | |
| Espécie | Taxa de Repetição
da DOE
Hz (25-27ºC) |
|
|
| 1. Hypopomus sp. 1a | 11 - 43 |
| 2. Hypopumus sp. 2 | 30 - 60 |
| 3. Hypopumus sp. 3 | 27 - 63 |
| 4. Hypopomus sp. 4 | 3 - 30 |
| 5. Hypopomus sp. 5 | 66 - 76 |
| 6. Hypopygus sp. b | 46 -62 |
|
|
| 7. Hypopomus sp. 9c | 30 - 60 |
| 8. Steatogenys elegans d | 50 - 60 |
| 9. Rhamphichthys rostratus e | 40 -100 |
|
|
| 10. Gymnohamphichthys sp. f | 25 -35 |
| 11. Gymnotus carapo
(Também # 8 e 9 acima.) |
45 -60 |
| ESPÉCIES PULSADORAS | |
|
|
| 12. Sternopygus macrurus | 60 - 150 |
| 13. Eigenmannia macrops | 430 - 660 |
| 14. Eigenmannia sp.
possivelmente E. troscheli ou Rhabdolichops sp. |
1580 - 1000 |
| 15. Eigenmannia sp.
2
(focinho pontudo) |
180 - 250 |
| 16. Adontosternarchus
sp.
(corpo manchado) |
1950 - 1250 |
| 17. Adontosternarchus
sp. 2
(corpo uniformemente cinza escuro) |
710 -1330 |
| 18. Apteronotus anas | 1080 - 1250 |
| 19. Sternarchella sp. | 1080 - 1170 |
| 20. Sternarchorhynchus oxyrhynchus | 1400 - 1800 |
| 21. Porotergus sp. (não foi gravada) | |
| 22. Sternarchorhamphus mülleri | 740 - g73 |
|
|
| 23. Sternarchella sp. | 934 -1384 |
| 24. Sternarchella sp. | ca. 2100 |
| 25. Sternarchogiton sp. | 889 -1241 |
| 26.Apteronotus sp.
(Também # 12, 13 (sp.;), 16, 20, 22.) |
900 - 1100 |
b) O mesmo que acima, com preferência para o folhedo e raizame denso; gregário, simpátrico com espécies acima, c) Substrato lamaçoso de margens com gramíneas inundadas, animais isolados, d) Margens com gramíneas inundadas, animais isolados, e) Margens com gramíneas inundadas, próximo ao fundo, animais isolados ou em pares, f) Essa é uma freqüência "perturbada e diurna". Valorores mais normais diurnos são 10-15, à noite 65-100 (Lissmann & Schwassmann, 1965). |
|
Fig. 9 - Eletrorreceptores. Representações esquemáticas de vários tipos de receptores bulbosos e tuberosos. A-H, as células ciliares modificadas ou células sensoriais secundárias. I-J órgãos do sentido inteiros mostrando a disposição das células sensoriais e sua inervação. A, ampola de Lorenzini dos elasmobrânquios; B-D, órgãos bulbosos dos teleósteos, incluindo gimnotóides; E, um tipo especial de órgão tuberoso em Mormyridae chamado "mormyromast" sc2; F, um órgão em Gymnarchidae chamado "gymnarchomast" II; G, "gymnarchomast" I; H, "mormyrimast" sc1; I, órgão bulboso em gimnotóide; J. órgão bulboso em gimnotóide. n, terminação nervosa mielinizada (terminações nervosas em I, pretas e J, brancas); sc, células sensoriais; camadas de pele indicadas por tipos diferentes de pontilhados (de Szabo, 1974).
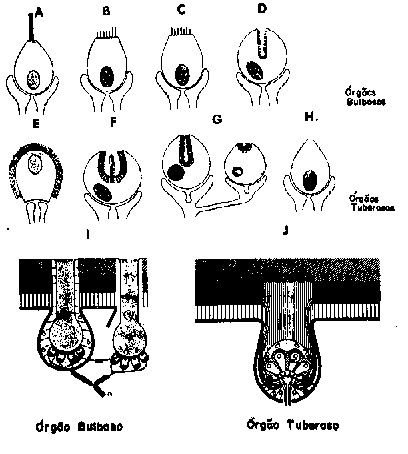
Em resumo, estes órgãos são
considerados como derivados especializados do sistema de receptores da
linha lateral, que normalmente respondem a correntes elétricas.
Essas correntes podem ser ou da própria DOE do peixe ou de outras
fontes animadas ou inanimadas. Eletrorreceptores existem não só
nos grupos mencionados acima, como também nos siluróides,
braquipterígeos (Polyodon, o "spoon bill" e Scaphirhynchus,
o esturjão) e provavelmente em alguns outros grupos.
Existem duas grandes classes e várias subclasses
destes órgãos de sentido, As grandes classes de eletrorreceptores
são (a) bulbosos ou sensíveis à baixa freqüência
e (b) tuberosos ou sensíveis à alta freqüência
(Fig. 9). Os primeiros existem em todos os grupos de peixes já citados,
os últimos ocorrem apenas nos teleósteos elétricos,
i.e., aqueles com órgãos receptores elétricos.
Fig. 10 - Uso da eletrorrecepção por um elasmobrânquio. Experimentos com um tubarão, Scyliorhimus canicula, mostram que a resposta ao alimento pode ser guiado por correntes elétricas fracas, de frequência muito baixa (cerca de 1 Hz) de um peixe presa (o "flatfish", Pleuronectes plateessa) ou de eletródios na areia (a) o mergulho normal, acuradamente dirigido a um "flatfish" enterrado na areia, b) disto persiste mesmo se o peixe for coberto por uma capa de agar eletricamente transparente. (c) o odor, quando exagerado pelo retalhamento da presa, leva a buscas não direcionadas rio abaixo (d) um filme plástico, eletricamente opaco, impede a resposta. (e) eletródios que simulam o campo bioelétrico de baixa frequência de um "flatfish" são atrativos, (f) eletródios (apenas um é mostrado) são localmente mais atraentes do que um pedaço de peixe (do Kalmljn, 1971).

Os receptores bulbosos são os mais sensíveis,
especialmente os das espécies marinhas, onde o forte curto-circuito
da água de alta condutibilidade torna o sinal .fraco. São
usados primariamente para detectar corrente contínua ou as lentas
mudanças nos campos elétricos da presa (Fig. 10) e das fontes
inanimadas. Uma destas é a corrente induzida pelo movimento das
massas de água, como as correntes marinhas, através do campo
magnético da terra, (Kalmijn, 1974). Estes receptores são
os mais sensíveis às correntes elétricas, variando
um equivalente a cerca de 1-10 Hz, dependendo da espécie.
Os receptores tuberosos são de pelo menos
3 ou 4 sub-classes, baseando-se no modo pelo qual eles codificam as diferenças
de intensidade. Provavelmente alguns são primariamente detectores
da DOE do próprio peixe, outros são usados primariamente
na eletrocomunicação - detectando as DOEs de outros membros
da mesma espécie. Os receptores tuberosos são os mais sensíveis
às flutuações das correntes elétricas de cerca
de 100-1.000 Hz, dependendo da espécie e mesmo do indivíduo.
Em espécies onduladoras como Eigenmannia
(DOE de cerca de 300 Hz), cada peixe tem sua própria frequência
de DOE normalmente preferida, dentro da variação da espécie,
que pode ser 200 Hz ou mais. Os receptores de um determinado peixe são
mais sensiveis às freqüências próximas a sua própria
taxa de DOE. A sensibilidade oscila tanto acima quanto abaixo desta "melhor
.frequência", i.e., os receptores são sintonizados, como filtros
passa-banda.
O cérebro desenvolveu sistemas receptores
e processadores especializados para a estimulação maciça
dos numerosos receptores. Estes são modificações
dos antigos centros da linha lateral. lncluem estruturas nos lobos da linha
lateral no bulbo, no cerebelo e no torus semicirculares do mesencéfalo.
Estas estruturas são extremamente grandes e altamente diferenciados
nos peixes gimnotoides e mormiriformes. As vias dos eletrorreceptores também.
se projetam no telencéfalo. O cerebelo é excepcionalmente
grande, não apenas nos peixes elétricos, mas também
nos elasmobrânquios, em geral, e em muitos siluroides. Existe uma
certo evidência de que isto esteja em conexão com a eletrorrecepção,
mas as informações são ainda insuficientes.
Muitos neurônios altamente especializados
existem nos lobos da linha lateral, no torus semicirculares e no cerebelo
dos gimnotóides e respondem aos sinais, derivados de correntes élétricas
e possuidores de padrões altamente específicos. Por exemplo,
um tipo de neurônio apenas responde à presença de um
outro peixe elétrico da mesma espécie, com uma taxa do DOE
que seja ligeiramente maior - não muito - do que a taxa de DOE do
próprio peixe (Fig: 11). No cerebelo, certos neurônios respondem
apenas quando a estimulação eletrorreceptiva vem de um objeto
que esteja se movendo em uma certa direção na água
próxima ao peixe. Assim, parece que eletrolocação
e a eletrocomunicação sejam servidas por sistemas distintos
de analisadores centrais que são responsáveis por aspectos
diferentes da estimulação do conjunto de receptores na pele.
Finalmente é suposto que o sistema da linha
laterel tenha.evoluido, entre seus resultantes especializados, uma parte
na códea dos vertebrados superiores. Assim, a pesquisa de
eletrorrecepção oferece uma oportunidade especial para desenvolver
uma abordagem comparativa ao estudo de nosso próprlo ouvido interno.
Incluindo o órgão auditivo. É impressionante que a
sintonia das fibras dos nervos aferentes dos eletrorreceptores tuberosos
em Eigenmannia é tão Seletiva Como a das. Fibras do nervo
coclear de mamíferos que tem a mesma frequência ótima,
embora sintonia deste seja considerada derivada principalmente das propriedades
mecânicas do ouvido.
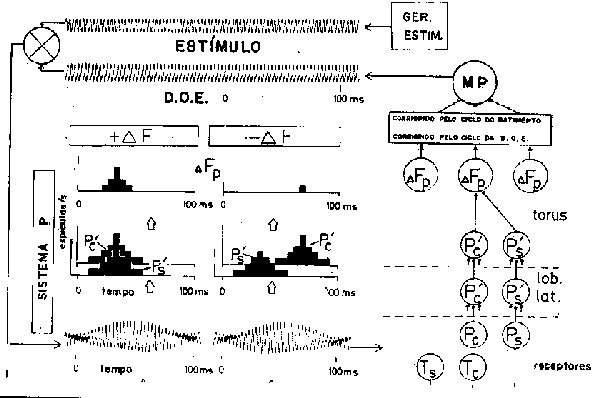
Fig. 11 - Processanento nervoso central da estimulação do eletrorreceptor. O diagrama mostra os componentes de um esquema dos passos sucessivos na Resposta de Esquiva Saltando (RES) em Eigenmannia. A descarga do órgão elétrico (DOE) é comandada pelo núcleo marca-passo (MP) fisiologicamente uma única unidade. O estímulo inicia a descarga de um vizinho Eigenmannia de uma frequência ligeiramente diferente, causando batimentos. Estas são de forma assimétrica porque a DOE não é sinusoidal; esta assimetria é essencial para a capacidade dos neurônios de distinguir corretamente entre +D F e -D F, i.e. se a frequência do vizinho é mais alta ou mais baixa e portanto a RES deve aumentar ou baixar. Os pequenos círculos representam tipos de neurônios geralmente encontrados nos níveis do sistema nervoso indicado (lob. lat. - lobus lateralis do bulbo; torus = torus semicircularis do mesencéfalo). Outros tipos estão sem dtivida envolvidos também e a figura mostra apenas uma metade do esquema. As fibras aferentes periféricas do tipo P têm uma maior prababilidade de disparar uma vez cadaciclo de DOE numa certa parte do ciclo de batimento. Esta parte é diferente para +D F e -D F em uma subclasse, as unidades Pc, mas não é diferente em outra subclasse, as unidades Ps, como mostrado no histograma de sua taxa de descarga. A convergência destes no torus explica a capacidade das unidades D Fp de assinalar o sinal do D F e controlar a direção de deslocamento do MP (de Scheich & Bullock, 1974).
AGRADECIMENTOS: A colaboração dos autores neste trabalho fui ajudada pelo Conselho Nacional do Desenvolvimento Científico e Tecno!ógico (CNPq), a Universidade Federal de Minas Gerais, a Deutsche Forschungsgemeinschaft, a Technische Hochschule, Darmstadt, a Universidade de Gottingen, a Universidade da California, San Diego, a Fundação Nacional de Ciências e o Instituto Nacional de Saúde, E.U.A. Os autores expressam seu agradecimento especial ao Dr. Warwick Kerr e ao pessoal do Instituto Nacional de Pesquisas da Amazônia, Manaus, especialmente aqueles do Departamento de Ictiologia, pela hospitalidade extremamente proveitosa.
| <<voltar |
PESQUISA > Peixe elétrico, caramujo e mamíferos > Link 11 > |