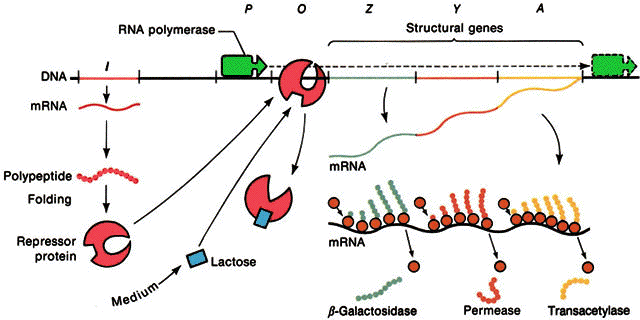
Controle da expressăo gęnica em procariontes
OPERON LAC
No metabolismo da lactose por E. coli săo necessárias duas enzimas que fazem parte de um operon chamado lac. A primeira enzima é a BETA-GALACTOSIDASE. Ela é capaz de clivar a lactose em glicose e galactose que assim servirăo como fonte de carbono para a célula (ver figura abaixo). A beta-galactosidase é dita uma enzima indutível, ou seja, sua expressăo varia com as necessidades celulares:
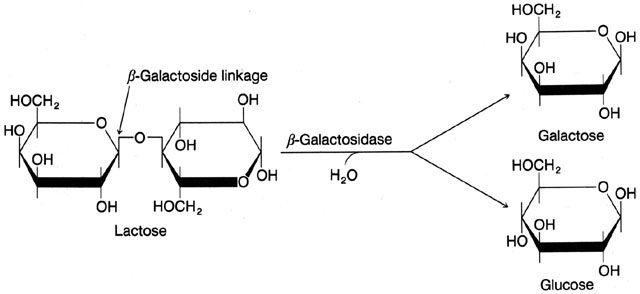
A outra enzima do operon é a PERMEASE, que, como seu próprio nome indica, é a enzima responsável pelo transporte de lactose do meio extracelular para o meio intracelular através da membrana bacteriana (a lactose, como a maioria dos carboidratos, năo é capaz de atravessar a bicamada lipidica sem uma proteína carreadora).
Existe ainda no operon lac uma outra enzima: Tiogalactosídeo Transacetilase. Seu papel in vivo ainda é incerto, mas in vitro é capaz de transferir uma acetila do acetil CoA para a hidroxila do carbono 6 de um tiogalactosídeo.
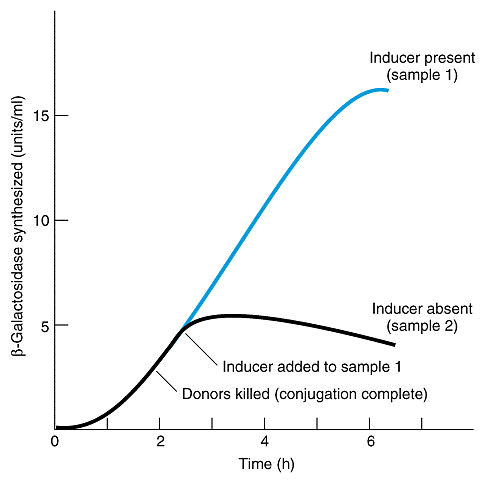
Um importante indício de que a induçăo ocorreria no metabolismo da lactose foi o aumento observado de beta-galactosidase quando se tinha um aumento de permease e tranacetilase.
Isto foi facilmente compreendido com modelos mutantes que mostravam que as 3 enzimas săo codificadas por 3 genes contíguos (ver figura abaixo), onde:
z - beta-galactosidase; y - permease; a - transacetilase
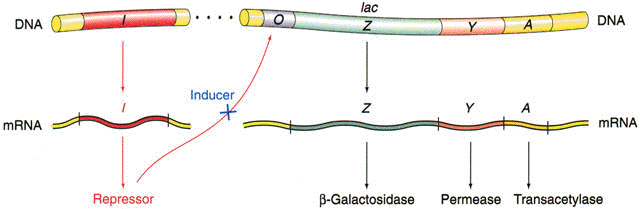
O RNAm do operon lac é dito policistrônico ou poligęnico pois possui informaçăo para codificar as 3 proteínas: beta-galactosidase, permease e transacetilase (rever figura acima).
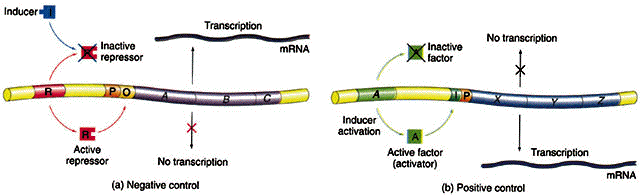
Depois da descoberta destes 3 genes, descobriu-se um mutante que possuia os genes para expressar as 3 proteínas, mas năo o fazia. Jacob e Monod deduziram que "a taxa de síntese destas proteínas é normalmente governada por um elemento comum diferente dos genes que especificam sua estrutura". A este gene deu-se o nome de regulador (gene i), sendo que os mutantes constitutivos possuem genótipo i-z+y+a+ e os selvagens i+z+y+a+.
O gene i é capaz de codificar um repressor que está faltando ou está inativado nos organismos com o gene i-.
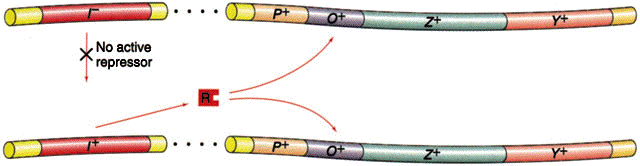
Através de estudos usando bactérias parcialmente diplóides, através do fator sexual (F'), observou-se que uma bactéria que possui o genoma i+z- e o fator F'i-z+ năo é capaz de metabolizar a lactose, ou seja, o repressor (produto do gene i) é difundível.
Existe no opreron lac uma regiăo promotora (p), que é o local onde a RNA polimerase se liga para começar a transcriçăo. Juntamente com o operador (o) formam os locais de controle do operon. Na presença do produto do gene i o operon é incapaz de ser codificado pois o repressor está ligado ao operador.
O isopropil-b-D-galactosídeo (IPTG) é um indutor artificial do operon lac. Foi através de estudos com esta substância que se descobriu o repressor lac, que na ausęncia do indutor liga-se ao operador e bloqueia a transcriçăo (veja as figuras abaixo).
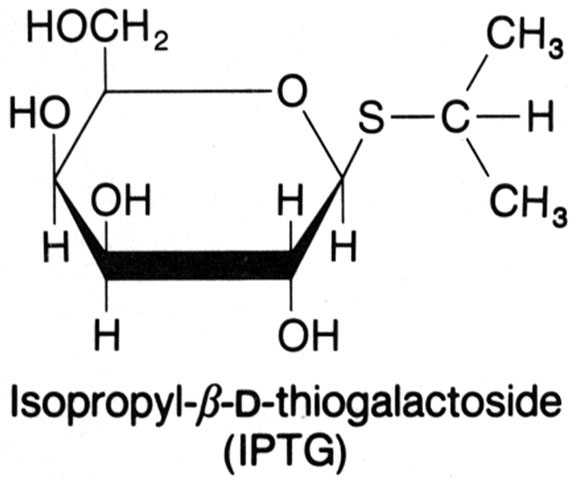
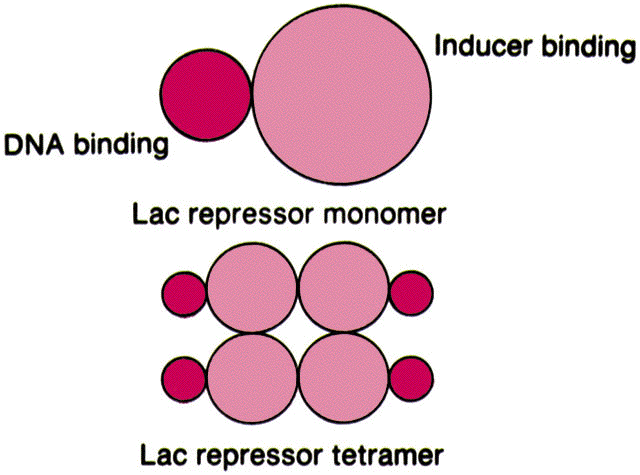
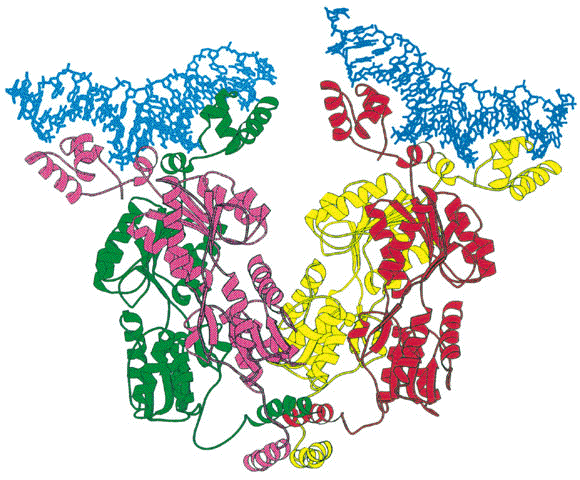
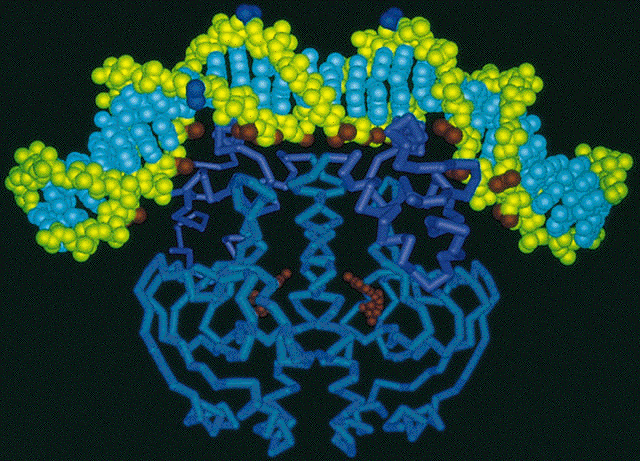
O repressor é uma proteína tetrâmera com subunidades idęnticas (de 37Kd), cada uma com um ponto de ligaçăo ao indutor. Através de cristalografia (ver fguras abaixo) descobriu-se que esta proteína possui eixos bilaterais, como seria esperado de uma proteína que se liga ao DNA. "O repressor encontra o operador se difundindo ao longo da molécula de DNA (uma procura unidimensional) e năo o encontrando a partir do meio aquoso (uma procura tridimensional".
Já é sabido que se a glicose estiver no meio de cultura da E coli, ela será preferencialmente usada como fonte de energia, isto é, enzimas utilizadas no metabolismo dos outros carboidratos serăo pouco ou nada expressas. Este efeito é chamado repressăo por catabólito.
Mas como se dá essa repressăo?
Um fato importante é que a glicose abaixa a concentraçăo do AMP cíclico em E coli. Através de estudos com AMPc exogeno, concluiu-se que "AMPc estimula o início da transcriçăo de muitos operons indutíveis" sendo, portanto, um sinalizador tanto em bactérias quanto em mamíferos.
Mas como se dá essa modulaçăo da transcriçăo via AMPc?
Em bactérias, o AMPc liga-se ao CAP (catabolite gene activator protein - proteína ativadora de genes por catabólitos), uma proteína dimérica com 2 subunidades idęnticas de 22Kd. Cada subunidade possui um domínio de ligaçăo ao DNA e ao AMPc (ver figura abaixo). Somente o complexo CAP-AMPc é capaz de estimular a transcriçăo e se ligar a determinados promotores (Saiba mais sobre motivos proteicos de ligaçăo ao DNA clicando AQUI!). No operon lac, CAP se liga próximo a a regiăo que se liga a RNA polimerase.
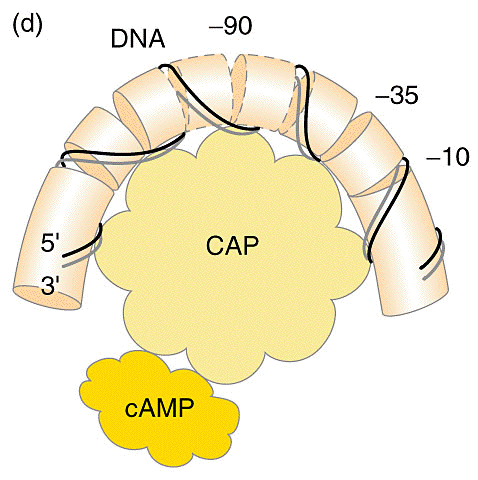
CAP exibe simetria bilateral que se ajusta ŕ seu ponto de ligaçăo do DNA. O repressor se liga em um regiăo "a frente" da RNA polimerase, portanto năo bloqueia a sua ligaçăo mas impede sua progressăo. CAP também estimula a transcriçăo do operon lac por possuir uma regiăo de ligaçăo a RNA polimerase quando está ligado ao DNA. Ele também é capaz de girar o DNA em 94 graus. Especificamente, 2 temas hélice-alça-hélice do dímero se inserem em sulcos maiores sucessivos da dupla hélice de DNA. Dois giros de 43 graus correspodem pela maior parte do giro para a direita do eixo do DNA. As ligaçőes do CAP e da RNA polimerase se reforçam mutualmente porque inclinam o DNA no mesmo sentido (uma visăo geral pode ser vista no modelo cristalográfico abaixo).
Os operons catabólitos indutíveis foram colocados sobre controle duplo, pois possuem promotores de ligaçăo acima da regiăo do operon (provavelmente enfraquecido durante a evoluçăo) para tornar operons dependentes de uma proteína auxiliar que inicia eficientemente a transcriçăo. O indutor específico atua em um só operon enquanto que o complexo CAP-AMPc é capaz de afetar muitos.
Operons controlados em conjunto pelo AMPc săo membros de um circuito regulador global, dentre eles: genes heat-shock (que săo ativados por uma subunidade sigma diferente da RNA polimerase quando há altas temperaturas) e genes SOS (que codificam enzimas de reparo após dano no DNA).
Uma visăo geral do operon lac pode ser feita na primeira figura dessa página
![]()
![]()
