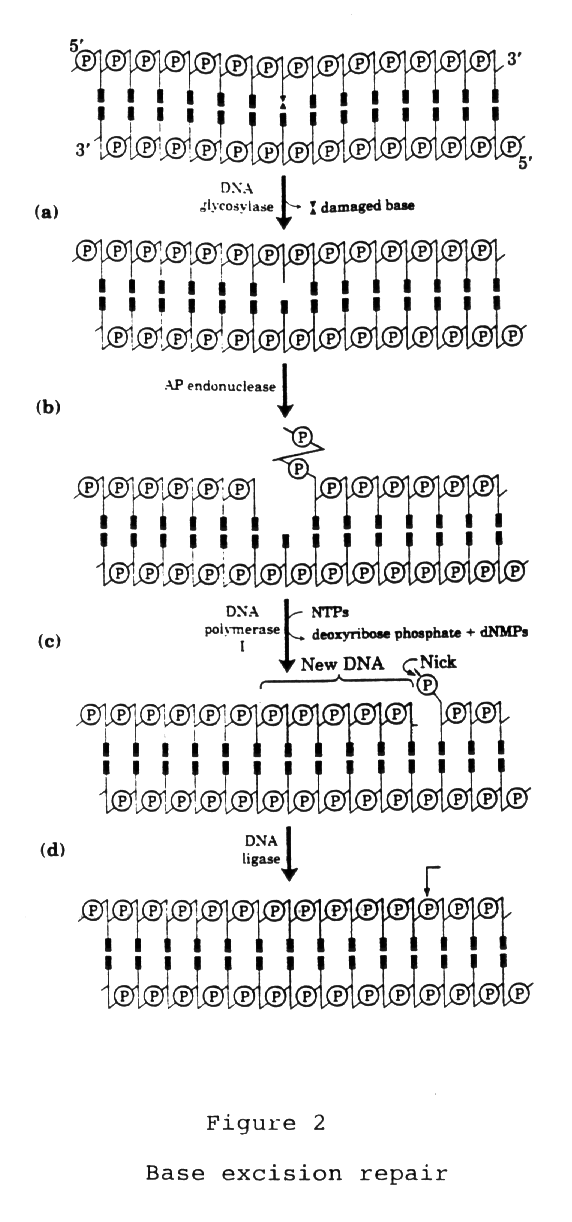
Este processo é conduzido pelas enzimas DNA glicosilases que
reconhecem os produtos de citosina e adenina deaminadas no DNA gerando
uracila e hipotanina. Esse tipo de
reparo tem início com a ação de DNA glicosilases que
reconhecem e removem bases lesadas da cadeia do DNA pela quebra da ligação
N-glicosil, a qual mantém a base nitrogenada associada com o esqueleto
de açúcar-fosfato (Friedberg et al., 1995). As enzimas com
atividade de DNA glicosilase foram inicialmente estudadas e caracterizadas
em E. coli, mas atualmente diferentes DNA glicosilases foram caracterizadas
em eucariotos e essas apresentam grande homologia com as enzimas procariotas.
A maioria das DNA glicosilases reconhecem lesões específicas
como a adenina metilada na posição 3 (Steinum & Seeberg,
1986) ou a 8-hidroxiguanina (Boiteux et al., 1987).
Após a retirada da base lesada gera-se no DNA um sítio apurínico
ou apirimidínico (AP) que será reparado por AP endonucleases
(Doetsch & Cunningham, 1990). Os sítios AP também podem
ser gerados por hidrólise natural do DNA. Todas as AP endonucleases
conhecidas até o momento realizam a quebra do esqueleto açúcar-fosfato
na região 5 deste. O resultado dessa quebra gera duas terminações:
uma 3-OH e outra 5-fosfato-desoxirribose.
A E. coli apresenta duas AP endonucleases: exonuclease III (xth) e endonuclease
IV (nfo). A exonuclease III fo i inicialmente caracterizada
como uma exonuclease com uma atividade associada de fosfatase (Doetsch
& Cunningham, 1990). Atualmente, sabe-se que a principal função
fisiológica dessa proteína é a atividade de 5 AP
endonuclease. Essa enzima tem aproximadamente 28 kDa e é capaz
de atuar eficientemente tanto em sítios apurínicos quanto
apirimidínicos (Gossard & Verly, 1978).
Por sua vez, a endonuclease IV de E. coli
não é detectada em extrato bruto de células selvagens,
uma vez que ela é responsável por somente 10% da atividade
de AP endonuclease em extratos brutos (Ljungquist, 1977). Essa enzima apresenta
atividade AP endonuclease semelhante à exonuclease III.
Após a ação das AP
endonucleases é necessário que ocorra a remoção
de resíduos 5-fosfato-desoxirribose. Essa etapa é realizada
pela enzima DNA desoxirribo-fosfodiesterase (dRpase), para que uma DNA
polimerase possa reconhecer e complementar a lacuna gerada pela retirada
do nucleotídeo (Friedberg et al., 1995).
A similaridade e conservação
entre os genes de eucariotos e procariotos é também funcional,
já que genes de eucariotos são capazes de complementar
bactérias deficientes em genes envolvidos no reparo por excisão
de base (Friedberg et al., 1995). A possibilidade dessa complementação
ocorrer advém do fato das enzimas que atuam nesse tipo de reparo
reconhecerem lesões específicas e não precisarem montar
um complexo enzimático para a retirada da base lesada. (Figura
2)
{kind=link}